棟壔妛尋媶強 惗柦婡擻壢妛尋媶僙儞僞乕
丂丂丂丂丂丂丂丂丂丂丂丂丂挻旝宍懺尋媶僠乕儉丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂
峏怴擔丂2019擭10寧1擔
暷丂懞丂尋丂媶丂幒
Research
尋媶攚宨
丂惗偒暔偺宍傗摦偒偑偍傕偟傠偄
丂丂惗偒暔偵偼偄傠偄傠側宍偺傕偺偑偁傝傑偡丅懱偺拞偵偼偝傑偞傑側婍姱丄慻怐偑偁傝傑偡丅偝傜偵嵶朎偺拞偵傕僆儖僈僱儔偑拋彉傪帩偭偰攝抲偟偰偄偨傝丄嵶朎崪奿偑旤偟偄峔憿傪嶌偭偰偄傞偙偲傕偁傝傑偡丅偦偟偰丄惗偒暔丄摿偵摦暔偼嵶朎儗儀儖偐傜屄懱儗儀儖偵帄傞傑偱丄摦偔偲偄偆偙偲偑摿挜偱偡丅巹偨偪偼揹巕尠旝嬀儗儀儖偐傜偺宍懺妛揑側僶僢僋僌儔僂儞僪傪帩偭偰偍傝丄摦暔偺摦揑側拋彉偺宍惉偵嫮偔庝偐傟偰偄傑偡丅摦揑側拋彉偺宍惉側偳偲尵偊偽丄暘巕偐傜偱偒偰偄傞嵶朎偑偳偺傛偆偵暘巕偺惈幙傪棙梡偟側偑傜摦揑側拋彉傪惗傒弌偡偺偐偲偄偆惗柦偺杮幙偵傕娭傢傞偙偲偱偡丅傑偨丄嬤擭偺嵞惗堛椕傊偺拲栚偱傕傢偐傞傛偆偵丄摨堦偺嵶朎偑廤傑傞偩偗偱偼婍姱偲偟偰偺婡擻偼敪婗偝傟偢丄暋悢偺庬椶偺嵶朎偑揔愗側宍懺宍惉傪峴偭偰弶傔偰婍姱偲偟偰婡擻偡傞傛偆偵側傞偺偱偡偐傜丄宍懺宍惉偺巇慻傒偑傢偐傞偙偲偼婎慴偐傜墳梡傑偱暆峀偔恖乆偺婜懸偵摎偊傞偙偲偵側傞偱偟傚偆丅
丂偳偆傗偭偰偦傟偑婲偙傞偺偐丠丂
丂丂偝傑偞傑側宍懺偑尒傜傟傞偙偲丄摦偒偑偁傞偙偲側偳偼偐側傝傛偔挷傋傜傟偰偒偰偄傑偡丅嵟嬤偱偼僎僲儉偑夝撉偝傟偨庬傕懡偔側傝丄儌僨儖惗暔偺曄堎懱偺夝愅摍偐傜傕宍懺宍惉偵廳梫側栶妱傪偡傞堚揱巕傕懡偔傢偐偭偰偒偰偄傞偲尵偊傞偱偟傚偆丅偟偐偟丄堚揱巕偺娭學偑柧傜偐偵側偭偨偩偗偱偼丄傢偐偭偨婥偵側傜側偄偺偑巹偨偪偱偡丅幚嵺偵嵶朎偺拞偱偳偺傛偆側偙偲偑婲偙偭偰偄偰丄偪傚偭偲傗偦偭偲偱偼棎傟側偄丄拋彉偩偭偨宍懺宍惉偑婲偒傞偺偐丅偦傟傪柧傜偐偵偡傞偺偑巹偨偪偺妛栤揑側嫽枴偱偡丅嵶朎偐傜嵶朎廤抍偺儗儀儖偱偺幚尡傪峴偭偰丄傾僀僨傾傪専徹偟偰偄偒傑偡丅柧夝側寢榑傪摼傞偨傔偺岻柇側幚尡宯傪偄偐偵慻傓偙偲偑偱偒傞偐丄崱傑偱偵抦傞偙偲偺偱偒側偐偭偨廳梫側忣曬傪摼傞偨傔偵偳偺傛偆偵媄弍傪摫擖偟偨傝奐敪偡傞偐偲偄偆偙偲偑偲偰傕廳梫偱丄尋媶幰偲偟偰偺僙儞僗偑忢偵帋偝傟傞尋媶暘栰偱偡丅丂
丂傛偔尒傞偙偲偐傜巒傔傛偆
丂丂巹偨偪偺僶僢僋僌儔僂儞僪偲偟偰丄岝妛尠旝嬀丄揹巕尠旝嬀儗儀儖偺幙偺崅偄娤嶡媄弍傪桳偡傞偙偲偑偁傝傑偡丅偙傟傜偺媄弍傪摉偨傝慜偵巊梡偡傞偲偄偆揰丄傑偨宱尡偵攟傢傟偨嵶朎憸偺夝庍傪巹偨偪偺傾僾儘乕僠偺拞偵嵟戝尷偵惗偐偡偙偲偑丄巹偨偪偺尋媶偺傗傝曽偺摿怓偱偡丅偦偟偰暘巕傪崻嫆偵偡傞棟夝偑昁梫偱偁傞偲峫偊偰偄傞偨傔丄堚揱巕敪尰丄僲僢僋僟僂儞側偳偺庤朄偺傎偐丄僞儞僷僋幙暘巕偺惈幙傪寁應偡傞偙偲傕峴偄傑偡丅愊嬌揑側嫟摨尋媶偱廳梫側暘巕偵偮偄偰偼X慄寢徎夝愅偵傛傝峔憿寛掕傪偟傑偡丅偦偺忣曬偐傜愴棯揑側曄堎傪摫擖偟偰暘巕儗儀儖偺抦尒傪怺傔傑偡丅偝傜偵嵟嬤偱偼椡妛揑僔僌僫儖偵拝栚偟丄偦偺宍懺宍惉偵偍偗傞悢棟揑側棟夝傕嫟摨尋媶偵傛傝巒傔偰偄傑偡丅
丂丂丂丂
尋媶僾儘僕僃僋僩
丂忋旂嵶朎偼丄敪惗偺嵟弶偵棏嵶朎偑暘楐偟偰巒傔偵偱偒偰偔傞嵶朎偱偡偟丄傑偨丄懱傪旐偄丄昞旂偐傜徚壔娗傑偱偝傑偞傑側宍懺宍惉傪峴偄側偑傜丄婡擻揑側婍姱傪嶌傞拞怱揑側嵶朎偱偡丅懱傪旐偆偲偄偆惈幙偐傜丄惗懱奜偲惗懱撪偲傪暘偗傞婎杮揑側栶妱傪帩偪傑偡丅摨庬偺嵶朎偑宷偑偭偰僔乕僩傪嶌傞偵偁偨偭偰丄忋旂嵶朎偼嵶朎娫愙拝憰抲傪嶌傝傑偡丅愙拝僞儞僷僋偲嵶朎娫傪椡妛揑偵宷偘傞偨傔偵嵶朎崪奿偲偺楢寢傪扴偆僞儞僷僋嶌偭偰偄傞偺偑傾僪傊儗儞僗僕儍儞僋僔儑儞(Adherens
junction, AJ)偲屇偽傟傞嵶朎娫愙拝憰抲偱偡丅忋旂嵶朎偱偼惗懱撪偺娐嫬傪堐帩偡傞偨傔丄暔幙偺桝憲傗偦傟傪巟偊傞僞儞僷僋丄僆儖僈僱儔偺攝抲偼曃偭偰偍傝丄偦偺曃傝偑偁傞偙偲傪忋旂嬌惈偲屇傃傑偡丅
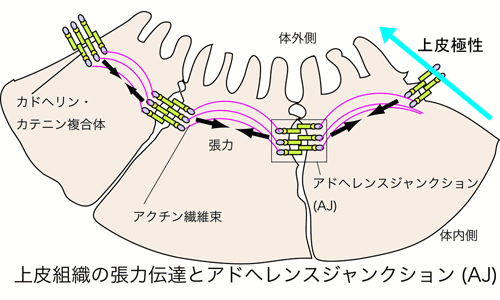
丂幚尡忦審傪帺桼偵曄偊堈偄攟梴嵶朎傪庡側嵽椏偵偟偰丄偱偒傞偩偗僔儞僾儖側幚尡宯偐傜婎杮揑偵廳梫側栤偄偵懳偡傞摎偊傪摼傛偆偲峫偊偰偄傑偡丅偦偺偨傔偵偼丄怴偨側傾僀僨傾偵傛傞幚尡宯偺妋棫傗丄媄弍奐敪側偳偺憂堄岺晇偑偲偰傕廳梫偱偡丅攟梴嵶朎偐傜婍姱宍惉丄屄懱傊偲偮側偑傞ES嵶朎偵偼丄戝偄偵拲栚偟偰偄傑偡丅
丒尰嵼偺僾儘僕僃僋僩
丂兛僇僥僯儞偺挘椡姶庴惈偲懡嵶朎宍懺宍惉偵偍偗傞堄媊
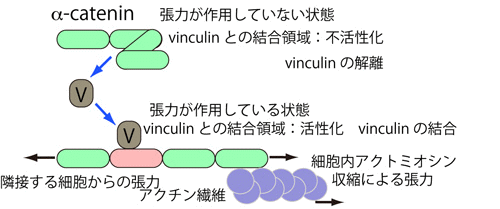
丂 忋偵弎傋偨嵶朎娫愙拝憰抲AJ偵偼傾僋僠儞慇堐偑寢崌偟偰偄傞偙偲偑抦傜傟偰偄傑偡丅傾僋僠儞慇堐偼儈僆僔儞慇堐偲憡屳嶌梡偟偰丄儌乕僞乕僞儞僷僋儈僆僔儞II偺椡傪揱偊丄偦偺寢壥丄AJ偼嵶朎偺撪懁偐傜堷偭挘傜傟傞偙偲偵側傝傑偡丅AJ偵偼嵶朎愙拝暘巕僇僪僿儕儞偑偁傝丄嵶朎偺奜偱偼椬愙偡傞嵶朎偺僇僪僿儕儞偲寢崌偡傞偨傔丄AJ傪夘偟偰嵶朎撪偺廂弅椡偑椬愙偡傞嵶朎偵揱傢傝傑偡丅偦傟偧傟偺嵶朎偱偦偺傛偆側偙偲偑婲偙傞偺偱丄堦斒偵忋旂嵶朎僔乕僩偺拞偱偼椬傝崌偆嵶朎偼AJ傪夘偟偰堷偭挘傝偁偭偰偄傑偡丅偦偺柧妋側棟桼偼柧傜偐偵側偭偰偄傑偣傫偑丄巹偨偪偼丄忋旂嵶朎僔乕僩偺堐帩偦偺傕偺偵廳梫偱偁傞偲峫偊偰偄傑偡丅嫮偄椡偑偐偐偭偰忋旂僔乕僩偑愜傟嬋偑傝丄恄宱娗摍傪嶌傞応崌偼傕偪傠傫丄AJ偵傛傞椡偺揔愗側揱払偑昁梫偱偡乮恾侾乯丅傑偨丄忋旂僔乕僩撪偺嵶朎偑巰傫偱丄忋旂偺僶儕傾婡擻偑棊偪偨応崌偵偼丄嵶朎偼偡偽傗偔丄巰嵶朎傪彍嫀偡傞偲偲傕偵偦偺応強傪椬愙偡傞嵶朎偑堎摦偡傞偙偲偵傛偭偰嵡偓傑偡丅偙偺偲偒傕丄嬌惈偺偁傞忋旂偺応崌偼丄AJ傪夘偟偰堷偭挘傝偁偆偙偲偱乮巰嵶朎偲惗嵶朎偺娫偱偼堷偭挘傝崌偄偼摉慠婲偙傜偢丄巰嵶朎偵柺偟偰偄傞惗嵶朎偺娫偱堷偭挘傝崌偄偑嫮偔側傞偲偄偆偙偲偑傢偐偭偰偄傞乯丄懝彎廋暅偑婲偙傞偺偱偡丅偙偺傛偆側応崌丄AJ偑偦偺憹壛偟偨廂弅椡偵懴偊傜傟偢偵夡傟偰偟傑偄丄嵶朎偑夝棧偟偰偟傑偭偰偼慡懱偺宍懺宍惉偵宷偑傝傑偣傫丅傑偨丄偦傟偧傟偺嵶朎偺廂弅椡偑姰慡偵摨堦偱偁傞偲偼峫偊傜傟偢丄懡彮偺嵎偑偁傞応崌丄嫮偄嵶朎偑廂弅偟丄廃埻偺嵶朎偑堷偭挘傜傟偰怢傃偰偟傑偆傛偆偱傕丄慡懱偺宍懺宍惉偵宷偑傝傑偣傫丅懡彮偺嵎偑偁偭偰傕丄堦恖彑偪傪婲偙偝偣側偄傛偆側丄巇慻傒偑偁傞偲岲搒崌偱偡丅
丂 巹偨偪偼AJ撪偺庡梫側傾僋僠儞寢崌僞儞僷僋偺堦偮價儞僉儏儕儞偑AJ偵椡偑偐偐傞帪偼廤愊偟丄椡偑偐偐傜側偔側傞偲夝棧偡傞偙偲傪尒偄偩偟傑偟偨(Miyake
et al. 2006)丅嫮偄椡偱堷偭挘傜傟偨帪偵偼偦傟偵懳峈偱偒傞傛偆偵傾僋僠儞寢崌僞儞僷僋偺検傪憹傗偟偰丄傛傝懡偔偺傾僋僠儞慇堐偑AJ偵寢崌偱偒傞傛偆偵偡傞偲偄偆巇慻傒偑偁傞偐偺傛偆偱偡丅椡偑摥偐側偔側傞偲價儞僉儏儕儞偼棧傟丄寢崌偡傞傾僋僠儞慇堐偺検偼尭傞偲峫偊傜傟傑偡丅廂弅椡偺崅偄嵶朎偑丄椬愙偡傞嵶朎傪堷偭挘傝偡偓偰丄AJ偑夡傟傞摍偟偨応崌偼丄傕偆椬愙偡傞嵶朎偑椡傪巟偊偰偔傟側偄偺偱丄椡偑偐偐傜側偄偙偲偵側傝丄價儞僉儏儕儞傕棧傟丄嵶朎撪偺廂弅椡偑AJ偵揱傢傜側偔側傝傑偡丅偙傟偼堦恖彑偪偟偦偆側嵶朎偑丄椡傪娚傔偰丄椬愙偡傞嵶朎偑堷偭挘傝曉偡偺傪懸偮偲偄偆偙偲偩偲峫偊傜傟丄偲偰傕偆傑偔偱偒偨僼傿乕僪僶僢僋偺婡峔偑偁傞偐偺傛偆偱偡丅
丂偦傟偼杮摉偐丄偳偺傛偆側巇慻傒側偺偐丄偦傟偑傢偐偭偰杮摉偺棟夝偵側傝傑偡丅價儞僉儏儕儞偑AJ偵傗偭偰偔傞偺偼丄僇僪僿儕儞偲暋崌懱傪嶌偭偰偄傞僞儞僷僋偺堦偮兛-僇僥僯儞偑寢崌憡庤偩偐傜偱偁傞偙偲偑偡偱偵傢偐偭偰偄傑偟偨丅偦偙偱丄兛-僇僥僯儞暘巕偵椡偺掱搙傪姶抦偟偰丄價儞僉儏儕儞偵寢崌偟偨傝偟側偔側偭偨傝偡傞巇慻傒偑偁傞偺偱偼丠偲峫偊傑偟偨丅偄傠偄傠偲幚尡傪偟偨偲偙傠丄恾俀偺傛偆偵丄兛-僇僥僯儞偵偼價儞僉儏儕儞偵寢崌偡傞椞堟偵椬愙偟偰丄偦偺寢崌傪幾杺偡傞慾奞椞堟偑偁傞偙偲偑傢偐傝傑偟偨丅偦偺慾奞偑夝偗傞偺偼丄兛-僇僥僯儞偑堷偭挘傜傟偨偲偒側偺偱偡丅偡側傢偪兛-僇僥僯儞偼挘椡姶庴惈傪帩偮AJ僞儞僷僋偱丄偦偺惈幙傪巊偭偰AJ偵傕挘椡姶庴惈乮椡偑偐偐傞偲價儞僉儏儕儞偑傗偭偰偒偰嫮壔偡傞乯惗傑傟偰偄傞傢偗偱偡(Yonemura
et al., 2010)丅
丂 尰嵼丄恑峴拞側偺偼丄偦傟偱偼兛-僇僥僯儞偺偳偙偑偳偆側偭偰慾奞偲偄偆偙偲偑婲偙傝丄偳偆側偭偰偦傟偑夝彍偝傟傞偺偐傪暘巕儗儀儖偱棟夝偡傞偙偲丄傑偨丄搒崌偺椙偄巇慻傒偩偲峫偊偰偒偰偄傑偡偑丄杮摉偵偳偺偔傜偄廳梫側偺偐傪丄摿偵忋旂嵶朎偺宍懺宍惉傪椺偵専徹偟偨偄偲偄偆偙偲偱偡丅忋旂嵶朎偼擇師尦攟梴偵尷傜偢嶰師尦攟梴傕峴偄丄惓忢側兛-僇僥僯儞傪敪尰偡傞嵶朎偲丄挘椡姶庴惈偑堎忢側兛-僇僥僯儞偺曄堎懱偺傒傪敪尰偡傞嵶朎偱偺堘偄傪尒偄偩偟偰偒偰偄傑偡丅峔憿惗暔妛偺愱栧壠傗悢棟壢妛偺愱栧壠偲嫤椡偟偰棟夝傪怺傔丄彨棃揑偵偼挘椡姶庴惈偑宍懺宍惉偵懳偟偰廳梫偱偁傞応崌傪尒嬌傔丄宍懺宍惉傪憖傞媄弍偺堦偮傊宷偘偨偄偲巚偄傑偡丅堦曽丄挘椡姶庴惈偑偦傟傎偳廳梫偱側偄応柺偱偼壗偑廳梫偱偁傞偺偐偲偄偆怴偟偄僥乕儅偑偁傇傝弌偝傟偰偔傞偲巚傢傟傑偡丅
丂兛僇僥僯儞偺傾僋僠儞寢崌惈偺挷愡婡峔偲偦偺堄媊
丂偙偺忋弎偟偨價儞僉儏儕儞寢崌偺挘椡姶庴惈偵娭學偡傞傕偺偺丄彮偟堘偆惈幙傪抦傠偆偲偟偰偄傑偡丅挘椡姶庴惈傪帩偮偲偄偆偙偲偼兛-僇僥僯儞偑堷偭挘傜傟傞偐傜偱偁傝傑偡偑丄偦偺N枛偱偼僇僪僿儕儞-僇僥僯儞暋崌懱傪宍惉偟傑偡偺偱丄椬愙偡傞嵶朎偐傜堷偭挘傜傟傑偡丅堦曽丄C枛偼傾僋僠儞慇堐寢崌擻偑偁傝丄嵶朎撪偺傾僋僠儞慇堐偵堷偭挘傜傟傞偲峫偊傜傟傑偡丅兛-僇僥僯儞偼僇僪僿儕儞-僇僥僯儞暋崌懱傪宍惉偟偰偄傞偲偒偼丄惗壔妛揑偵偼傾僋僠儞慇堐寢崌擻偑尒傜傟偢丄惛惢偝傟偨扨懱偱偼傾僋僠儞慇堐偵寢崌偡傞偙偲偑曬崘偝傟偰偍傝丄偦傟偐傜偡傞偲彮側偔偲傕兛-僇僥僯儞偺傾僋僠儞慇堐寢崌擻偼壗傜偐偺惂屼傪庴偗偰偄傞偲峫偊傜傟傑偡丅偟偐偟丄嵶朎撪偱偼AJ偵挘椡姶庴惈傪帵偡丄偡側傢偪丄僇僪僿儕儞-僇僥僯儞暋崌懱撪偺堷偭挘傜傟偆傞兛-僇僥僯儞偑懚嵼偟偰偄傞偙偲傕柧傜偐偱偡丅偲偄偆偙偲偼嵶朎撪偱傕忬嫷偵傛傝傾僋僠儞慇堐寢崌擻偼忋偑偭偨傝壓偑偭偨傝偟偰偄傞傛偆偵巚傢傟傑偡丅
丂埲忋偺偙偲偐傜丄傾僋僠儞慇堐偲兛-僇僥僯儞偲偺寢崌偺惂屼偺巇慻傒傕廳梫偱偍傕偟傠偄僥乕儅偩偲峫偊傜傟丄惛椡揑偵庢傝慻傫偱偄傞偲偙傠偱偡丅巹偼丄兛-僇僥僯儞偲傾僋僠儞慇堐偲偺寢崌偼帺揮幵偺僊傾偲僠僃乕儞偺娭學偺條偵丄堦曽曽岦偵椡傪偐偗傞帪偵偼姎傒崌偄丄偟偭偐傝寢崌偡傞偑丄椡偑偐偐傜側偄丄偁傞偄偼堎側傞曽岦偵椡偑偐偐傞帪偵偼姎傒崌偢丄寢崌偑尒傜傟側偄丄側偳偺巇慻傒傕偁傞偺偱偼側偄偐丄偲憐憸偟偰偄傑偡乮Yonemura
2011-a, Yonemura 2011-b乯丅
丂忋旂嵶朎偺嬌惈宍惉婡峔
丂忋旂嵶朎偼擇師尦偺僔乕僩傪嶌傞偺偑婎杮偱偡丅偦傟偑棫懱揑偵戃傪嶌偭偨偺偑懡嵶朎摦暔偺尨宆偱偡丅偙偺僔乕僩偺堄枴偼丄惗懱奜偲嬫暿偟偰惗懱撪偲偄偆娐嫬傪嶌傞偙偲偱偡丅忋旂僔乕僩偺惗懱撪懁偵偼婎掙枌偲偄偆摿庩側嵶朎奜婎幙(ECM)惉暘偐傜側傞峔憿偑愙偟偰偄傑偡丅忋旂嵶朎偺婎掙枌偵愙偡傞嵶朎枌傪basal
membrane偲屇傃丄偦偺斀懳懁丄惗懱偺奜偵柺偟偰偄傞嵶朎枌偼apical membrane偲屇傃傑偡丅椬愙偡傞嵶朎偲愙偟偰偄傞嵶朎枌偼lateral
membrane偱偡丅Basal membrane偲lateral membrane偲偼惈幙揑偵傎偲傫偳摨偠偲傕巚傢傟丄嬌惈傪峫偊傞忋偱偼basolateral
membrane偲丄偟偽偟偽堦偔偔傝偵偝傟傑偡丅塰梴摍傪媧廂偡傞忋旂嵶朎偼apical membrane偵偼塰梴偺僩儔儞僗億乕僞乕側偳偑攝抲偝傟偰偄傑偡丅偙偺応崌丄Apical
membrane偼偦偺柺愊傪戝偒偔偡傞偨傔偵丄旝鉕栄偺傛偆側傾僋僠儞慇堐偺懇傪幉偲偟偨摿庩側嵶朎崪奿偵棤懪偪偝傟偨宍忬傪庢偭偰偄傑偡丅庢傝崬傑傟偨塰梴偼媡栠傝偣偢丄basal
membrane偐傜曻弌偝傟傑偡丅傑偨丄lateral membrane偺apical傛傝偵偼AJ偲偝傜偵apical傛傝偵嬤愙偟偰tight
junction (TJ)偑偁傝傑偡丅摿偵TJ偼嵶朎娫偺暘巕偺堏摦乮楻傟乯傪惂屼偡傞峔憿偱偡丅忋旂僔乕僩偱暍傢傟偨嬫夋偺暘巕慻惉偺堘偄傪曐偮偨傔偵丄暘巕偺捠峴傪尩偟偔惂尷偡傞応崌傕偁傝傑偡丅偙傟傪忋旂僔乕僩偺僶儕傾婡擻偲尵偄傑偡丅傑偨丄TJ偼傓偟傠摿掕偺暘巕傪慖戰揑偵捠夁偝偣傞僠儍儞僱儖偺栶妱傪壥偨偡偙偲傕偁傝傑偡丅塰梴傪庢傝崬傓偨傔偵昁梫側僫僩儕僂儉僀僆儞偑惗懱奜偐傜丄惗懱撪偵擖偭偰偟傑偆偲丄嵶朎偼僫僩儕僂儉億儞僾傪巊偭偰僫僩儕僂儉傪basolateral懁偵媯傒弌偟傑偡丅偦偺傑傑偱偼apical懁偱偁傞娗峯撪偺僫僩儕僂儉擹搙偑壓偑傝塰梴偑媧廂偱偒側偔側傞偺偱TJ偑僫僩儕僂儉傪慖戰揑偵摟夁偝偣丄娗峯懁偵栠偟傑偡丅偙偺傛偆偵apical
membrane偲basolateral membrane偲偼暘巕慻惉傕峔憿傕婡擻傕堎側傝丄傑偨嵶朎幙偱傕桝憲偺曽岦惈傗嵶朎崪奿偺暘晍側偳暘巕丄峔憿儗儀儖偺曃傝偑偁傝傑偡丅偙傟傪忋旂嬌惈偲偄偄傑偡丅TJ偲AJ偲偼偪傚偆偳apical
membrane偲basolateral membrane偲偺嫬奅偵宍惉偝傟丄TJ偺婡擻傪峫偊偰傕忋旂嬌惈偲枾愙側娭學偑偁傞偲峫偊傜傟偰偒偰偄傑偡丅忋旂嬌惈偼忋偵弎傋偨傛偆偵懡嵶朎摦暔偺懱傪嶌傞忋偱杮幙揑偵戝愗側傕偺偱偁傝丄偦偺攋抅偼忋旂僔乕僩偺婡擻偺攋抅偵偮側偑傝丄嵶朎偺暘壔傗偑傫壔偲傕娭學偟偰偄傑偡丅偦偺偨傔忋旂嬌惈傪嶌傝丄堐帩偡傞婡峔偼挿偄娫拲栚傪梺傃懕偗偰偄傑偡丅娭學偺偁傞堚揱巕傕懡偔傢偐偭偰偒傑偟偨偑丄偦傟傜憡屳偺娭學傗apical偺惃椡傪峀偘傞場巕偱偁傞偲偐丄basolateral
偺惃椡傪峀偘傞場巕偱偁傞偲偄偆傛偆側懆偊傜傟曽傪偟偰偄偰慡懱憸偼偮偐傔偰偄傑偣傫丅弶傔偵壗傪棅傝偵丄壗偑嬌惈傪寛傔傞偺偐丄傑偨丄寛掕偟偨嬌惈偵廬偭偰幚嵺偺暘巕傗峔憿偺曃傝傪傕偨傜偡偺偼偳傫側巇慻傒偵傛傞偺偐側偳偼丄惍慠偲棟夝偝傟偰偄傑偣傫丅忋旂嬌惈偼忋旂偑僔乕僩偵側偭偨帪偵廳梫側偺偩偐傜丄嵶朎娫愙拝偲枾愙偵娭學偑偁傞偲挿傜偔怣偠傜傟偰偒傑偟偨偑丄偙偺侾侽擭偺偁偄偩偵丄嵶朎娫愙拝偑側偔偰傕偙偙偺忋旂嵶朎偼忋旂嬌惈傪嶌傝偆傞椺偑偄偔偮傕曬崘偝傟偰偒傑偟偨丅TJ側偳偺愙拝憰抲偑偱偒偨偙偲傪丄忋旂嬌惈偑偱偒偨徹嫆偲傒側偡尋媶幰傕傑偩懡偄偺偱偡偑丄偦偆峫偊傞偲丄忋旂嬌惈偵廳梫側場巕傕丄愙拝憰抲宍惉偩偗偵娭傢傞場巕傕尒暘偗偑偮偐側偔側傝傑偡丅偦偺傛偆側偙偲偐傜丄偟偭偐傝偟偨慡懱憸傪攃埇偡傞偨傔偵嵶朎娫愙拝傪婲偙偝偢丄偟偭偐傝偟偨忋旂嬌惈傪帩偮攟梴嵶朎傪巊偆偲偄偆丄岺晇傪偟偨宯傪巊偭偰丄忋旂嬌惈偺婡峔傪尋媶偟偰偄傑偡丅
丂忋旂嵶朎偵偍偗傞椬愙偡傞嵶朎偺惗巰傪擣幆偡傞婡峔
丂偙傟傕傑偨丄旕忢偵柺敀偄偗傟偳側偐側偐擄偟偄僥乕儅偱偡丅忋旂僔乕僩撪偺嵶朎偑巰偸偲偦偺嵶朎偼墴偟弌偝傟丄摨帪偵偦偺応強偑廃埻偺嵶朎偵傛偭偰嵡偑傟丄忋旂僔乕僩偺僶儕傾婡擻偺攋抅偑嵟抁偺帪娫偵側傞傛偆偵懝彎廋暅偑峴傢傟傑偡丅偦偺嵺偵偼巰嵶朎傪庢傝姫偄偰偄傞惗嵶朎偺巰嵶朎偵柺偟偨懁偵偺傒丄儈僆僔儞II偺廤愊偑尒傜傟傑偡丅偦偺儈僆僔儞II偼AJ偵寢崌偟偨傾僋僠儞慇堐偲憡屳嶌梡偟丄儈僆僔儞II偺摦偒偑AJ傪堷偭挘傞偙偲偵側傝傑偡丅偪傚偆偳巰嵶朎傪庢傝姫偄偰傾僋僩儈僆僔儞偑抐懕揑偵宍惉偝傟丄偦傟偑AJ偵傛偭偰宷偑傝堦偮偺娐傪宍惉偡傞偙偲偵側傝傑偡丅傾僋僩儈僆僔儞偺廂弅偵傛傝偦偺娐偼彫偝偔側傝丄偦傟偑懝彎廋暅傪彆偗傑偡丅偙傟偑偲偰傕柺敀偄偺偼丄惗嵶朎偼椬偺嵶朎偑惗偒偰偄傞偐巰傫偱偄傞偐傪慺憗偔敾抐偟丄巰傫偱偄傞嵶朎懁偵儈僆僔儞II傪廤愊偝偣傞偲偄偆偙偲偱偡丅妋偐偵丄惗偒偰偄傞嵶朎偺廃埻偵儈僆僔儞II傪廤愊偝偣丄惗偒偰偄傞嵶朎傪捛偄弌偟偰偟傑偭偰偼傑偢偄偟丄偦偺傛偆側偙偲偼婲偒傑偣傫丅忋旂僔乕僩偑寬慡偵堐帩偝傟丄偄偞攋抅偑婲偒傟偽丄嵟懍偱廋暅傪峴偆丄偦偺巇慻傒偑偙偙偵偁傞偼偢偱偡丅嵟傕娞怱側偺偼丄暔幙偲偟偰壗傪姶抦偟偰惗偲巰偲傪尒暘偗傞偺偐偲偄偆偙偲偱偡丅嵟嬤丄偙傟偑廳梫偩丄偲曬崘傪偟偰偄傞尋媶幰傕偄傞偺偱偡偑丄捛帋傕偱偒側偄偺偱丄巹偼昁偢偟傕慡柺揑偵偼怣偠偰偄傑偣傫丅偟偐偟側偑傜丄偦傟偱偼壗側偺偐丄偲偄偆偲側偐側偐撍攋岥偑奐偗側偄丄偦偺傛偆側僥乕儅偱偡丅
偙傟傑偱偺僾儘僕僃僋僩
僶僫乕僗儁乕僗
挻旝宍懺尋媶僠乕儉
仹650-0047
暫屔導恄屗巗拞墰嬫峘搰撿挰2-2-3
TEL 078-306-3105
FAX 078-306-3107